Introduction
Pharmacology
Ayahuasca is a beverage extracted from plants in the Amazonian rain forest. By boiling the bark of the vine Banisteriopsis caapi with the leaves of the plant Psychotria viridis a psychoactive brew is made. The Psychotria viridis contains the short-acting psychoactive agent dimethyltryptamine (DMT) which has a structural similarity to serotonin. When DMT binds to neuronal receptors in the brain it causes changes in cognition and state of consciousness. However, DMT is orally active only in the presence of MAO inhibitors, such as the beta-carbolines of the Banesteriopsis bark. Thus, the interaction between the DMT and beta-carbolines is the basis of the psychoactivity of ayahuasca (McKenna et al., 1984).
Ayahuasca has a long history of safe and beneficial use for spiritual and healing purposes among large numbers of indigenous people in South America (Metzner, R., 1999). Only recently has Western science become interested in studying ayahuasca scientifically. In 1993, the Hoasca Project, a multinational, biomedical study of the ayahuasca tea, took place in the Brazilian Amazon Basin and the tea's pharmacological, physiological and psychological effects on humans were studied (McKenna, D.J. et al., 1998; Callaway, J.C., et al., 1999; Grob, C.S., et al., 1996).
Electroencephalography (EEG)
There is general consensus that rhythmical waves recorded from the surface of the scalp (the EEG) are summed synaptic potentials generated by the pyramidal cells in the cerebral cortex. The EEG represents responses of cortical neurons to rhythmic discharges from the thalamus. The frequency and amplitude of the EEG are determined by a complex arrangement of excitatory and inhibitory interconnections within the thalamus itself and between the thalamus and the cerebral cortex. The spontaneous fluctuations of the brain electrical activity (EEG), picked up on the scalp surface by electrodes, vary from moment to moment in both amplitude (voltage) and frequency (cycles per second). In order to interpret this complex signal, quantification, usually by a computer, is necessary.
The Quantitative EEG (qEEG)
In order to assess the effects of a substance on brain functions, the quantitative EEG (qEEG) is useful. Usually mean amplitude or power within the major frequency bands is computed. Other variables such as right/left amplitude ratios and phase coherence between pairs of symmetrical EEG signals may be calculated. Ample evidence suggests that the qEEG is a valid measure of the functional state of the brain. Research has shown that there is a direct correlation between a person's level of conscious attention and his brain wave frequency. During the unconscious state of deep sleep, high amplitude, slow delta waves (0.5-4Hz) predominate the EEG. In the semi-conscious state between wakefulness and sleep (the drowsy, hypnagogic state) theta waves (4-8Hz) tend to prevail. During the awake, mentally relaxed state with eyes closed, alpha waves (8-13Hz) predominate the EEG in most people while the fast beta waves (13-30Hz), normally with smaller amplitudes, are observed during aroused, hyperattentive states of consciousness (Fig. 1).
Thus, the qEEG method can determine whether a person is conscious (alpha and beta waves), subconscious (theta waves) or unconscious (delta waves). The general rule is that the lower the EEG frequency the more unconscious the individual.

EEG research of psychedelics
The majority of EEG studies done on psychedelics appeared in the scientific journals some 30 years ago, before these compounds were banned. Wikler (1954), Itil (1968) and Fink (1978) are all in agreement that psychedelics, regardless of the substance (LSD, mescaline, psilocybin), produce decreases in slow wave (alpha and theta) activity together with increases of fast (beta) activity. This low amplitude, desynchronized EEG pattern induced by psychedelics reflects an activation of the brain and is in opposition to the highly synchronized alpha pattern observed during deep relaxation. Fink (1978) found that regardless of the nature of the drug administered, EEG synchronization (alpha/theta waves) was associated with euphoria, relaxation, and drowsiness; while EEG desynchronization was associated with anxiety, hallucinations, fantasies, and illusions. Don et al. (1998) found an increase of high frequency beta ('40Hz') with no significant change of alpha and theta activity in the EEG following the ingestion of ayahuasca. All the above studies indicate that most psychedelic compounds tend to suppress low EEG frequency activity (alpha and theta) and enhance beta activity reflecting an activation of the brain. However, other psychedelic-like compounds such as marihuana and MDMA (ecstasy) seem to have the opposite effect and increase alpha activity. In a recent, controlled placebo study, an increase of EEG alpha power, correlating with intense euphoria, was found after smoking marihuana (Lukas, et al., 1995).
Long-term effects of the use of psychedelics, using qEEG monitoring, have rarely been studied. However, in a recent study of 23 recreational MDMA users, Dafters et al. (1999) found that the use of MDMA was positively correlated with absolute power in the alpha (8-12Hz) and beta (12-20Hz) frequency bands. These findings were supported recently in another study by Gamma et al. (2000) who found global increases of theta, alpha and beta power in a group of regular MDMA users compared to a control group.
Consciousness expanding properties of ayahuasca
The main psychoactive agent in ayahuasca (Psychotria viridis) is the short-acting compound dimethyltryptamine (DMT). The only published study of the qEEG effects of DMT was done on mice with implanted electrodes (Morley BJ, Bradley RJ, 1977). These authors found a dose-dependent hypersynchrony with increased power in the low frequency (delta/theta) range following the ingestion of DMT.
DMT, when applied intravenously in humans, almost instantaneously elicits visual hallucinations, bodily dissociations and extreme shifts in mood (Strassman et al., 1994). It is the mind-altering properties of ayahuasca which have been used for healing and spiritual purposes in shamanic ayahuasca rituals by indigenous people throughout South America for maybe a thousand years (Metzner, R. et al, 1999).
In many respects, the changes in consciousness induced by drinking ayahuasca are comparable to other states of deep relaxation and increased subconscious awareness, such as the meditative, the hypnotic and the hypnagogic states which are all characterized by increased theta activity in the EEG (Hoffmann E., 1998; Ray WJ, 1997; Budzynski TH, 1986). Thus we expected to see increased theta and possibly increased alpha activity following the intake of ayahuasca.
Materials and Methods
Subjects
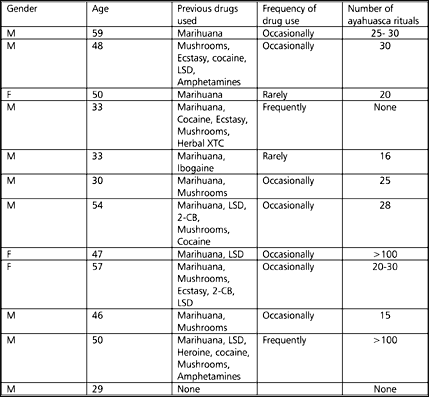
Subjects were twelve normal volunteers, mostly Europeans, who attended a workshop of four ayahuasca rituals in Brazil, organized by an experienced Ayahuasceros, a native healer (YSB), working already for years with ayahuasca. Subjects included nine men and three women between the age of 29 and 59 years (mean = 44.7 years). Two subjects had never taken ayahuasca before while the remaining ten had attended 12 to over a 100 ayahuasca rituals before. Most of the subjects had taken psychedelics as well as other drugs before (Table 1).
| Table 1. Demographic data of the subjects | ||
| ||
| At the time of the experiment none of the volunteers had any physical complaints. One male subject described himself as a first-class junkie, but refrained from drug-use, apart from marihuana and ayahuasca, since he started using ayahuasca in ritual settings. A second male subject suffered from decompensatio cordis and used prescription medication (digoxine, enalapril, and a diuretic). There was one male subject with a complicated psychiatric history, suffering from depressive symptoms in the past and was treated with serotonin uptake inhibitors. Most volunteers had a higher education or university degrees (law, medicine, psychology). | ||
In order to screen for psychiatric symptoms, the subjects initially underwent a psychiatric interview and rated themselves on the SCL-90 Symptoms Checklist. All subjects had scores within the normal range. All volunteers signed an extensive Informed Consent Form explaining the effects and side effects of ayahuasca, as well as the EEG recording procedure. It should be noted, however, that the intention of the volunteers was to experience drinking ayahuasca under the guidance of (YSB). The monitoring of the qEEG was the scientific part of the workshop; the workshop was explicitly not organized to monitor the qEEG.
The EEG data reported here were all collected under field conditions, in the middle of the Brazilian jungle, using batteries as the power source for the equipment. Baseline EEG recordings were done, together with the psychological testing, in the village of Alto Paraiso, north of the capital Brasilia. The rituals were held a few days later in a remote area, only accessible by foot.
Administration of ayahuasca
EEG recordings were done at two separate rituals led by YSB according to methods developed within the ethno-psychopharmacological society �Friends of the Forest' in the Netherlands and based on Brazilian shamanistic tradition. Three doses of ayahuasca, 75 ml in total, were administered during the ritual with intervals of 1-1 1/2 hours. EEGs were taken one to two hours following the third dose of ayahuasca, 4-6 hours after the first dose. At this point all subjects appeared to be experiencing an altered state of consciousness.
EEG equipment and electrodes
The PCEEG81, an eight channel EEG brainmapping instrument from Khepri Electronics, was used for data acquisition. This battery-operated portable unit, which weighs only 8 ounces, works with a laptop PC and is very suitable for field work.
Using 128 samples per second, 12 bit A/D conversion, eight channels of EEG and one channel of EMG are fed, via fiber optic transmission cable, to the serial port of a Pentium 233, PC laptop. The signals are simultaneously stored on a disk and displayed on the computer screen. Later, data may be replayed from the hard disk, edited for artefacts, and analyzed for frequency, amplitude and a number of statistical parameters.
We used bipolar scalp recordings from eight different electrode locations using the international 10/20 system. Gold-plated electrodes were fastened to the subject's head with elastic head bands, and EEG signals were obtained from right and left frontal, temporal, post-temporal and occipital lobes (F7, F8, T3, T4, T5, T6, O1, O2) with Cz on the top of the head as a common reference.
Data collection, editing and analysis
Baseline recordings were done in a hotel room in the village of Alto Paraiso. The subjects laid down on a couch with closed eyes while their EEGs were recorded. Following a few minutes of rest, approximately 200 seconds of EEG signals were stored on disk for later analysis.
A few days later all the subjects had their EEGs recorded again during two separate ayahuasca rituals in the jungle. By the end of the ritual most people were lying down quietly absorbed in their inner subjective experiences. In order not to disturb the subjects and interfere with their state of consciousness, we did not want to take them to an adjacent room for the EEG measurements but decided to do the recordings in the very same spot in the Zendo (meditation hall) where they had been sitting or lying during the ritual. They did not even have to get up from their mattresses for the EEG. Thus we expected them to remain in an altered state of consciousness throughout the recordings. During the measurements the subjects were lying flat on their backs with closed eyes. The room was relatively quiet at that point, and all subjects seemed to be deeply relaxed and absorbed in their inner experiences. Thus, very few muscle and eye movement artefacts were observed in the recordings. Also electrical noise interference with the recordings was at a minimum since all our equipment was battery operated.
After the rituals the EEG records were replayed, inspected visually, and edited for eye/head movements and muscle tension artefacts. Power spectral analyses were done of at least 100 seconds of artefact-free EEG epochs. For each of the 8 scalp sites, the mean amplitude in the theta (4-8Hz), alpha (8-13Hz), and beta (13-20Hz) frequency bands were computed using Fast Fourier Transform (FFT) analysis. Since it is sometimes difficult to differentiate between delta waves and eye movement artefacts, we chose not to analyse delta (0.5-4Hz) frequencies in order to avoid misinterpretations.
Results
To assess the effects of ayahuasca on the resting EEG, mean theta, alpha and beta amplitudes following the ritual were compared to baseline values. Because the data could not be considered normally distributed, a non-parametric statistical test, the Wilcoxon Signed Ranks, two-tailed test, was used for comparative analysis.
We found considerable and statistically significant increases (p<.01 or p<.05) of mean alpha amplitudes after ayahuasca for the O1, O2 and T6 locations. For the left occipital lobe the increase of the group mean was an impressive 120%, statistically significant at p<0.01. Smaller but statistically significant increases of mean theta amplitudes were found at F7, F8, T3, T5, O1 and O2 locations. Eleven out of 12 subjects showed substantial increases of theta activity. Thus, increases of alpha amplitudes were seen primarily in the occipital regions of the brain and in the left post-temporal area, while increases of theta amplitudes were observed in all parts of the brain, except for the right temporal and post-temporal areas. Only one subject who initially had a low amplitude, inhibited EEG with virtually no alpha, showed almost no changes of his EEG under the influence of ayahuasca.
Discussion
We found large and statistically significant increases of both EEG alpha and theta mean amplitudes in a group of normal subjects following three doses of ayahuasca in a shamanistic ritual setting. To our knowledge this is the first study to demonstrate such profound EEG changes after the intake of ayahuasca. The only other published EEG study of ayahuasca (Don et al., 1998) found statistically significant increases only of high frequency (36-44Hz) beta, following the intake of ayahuasca. It's important to notice that these authors did their EEG recordings after only one dose of ayahuasca while we did our recordings following three doses of the tea. Also Don and collaborators took their subjects out of the ritual context to an adjacent room for EEG recordings and thus may have interfered with their altered states of consciousness. In addition, Don et al. found a trend toward decreases of alpha and theta power and an increase of 14-30Hz beta power. Thus, their subjects showed a somewhat activated EEG pattern which could be due to some apprehension following the intake of ayahuasca. On the contrary, our findings of increased alpha and theta amplitudes and unchanged beta activity point to the fact that our subjects were extremely relaxed and in an altered state of consciousness during the EEG recordings by the end of the ritual. This was also confirmed by the subjects during questioning after the recordings.
One may speculate that the increase of theta waves during the ritual was due to drowsiness in the subjects. Theta waves tend to prevail in the EEG during drowsiness and light sleep, while alpha waves usually decrease. Alpha waves are indicators of conscious attention, and since nearly all of our subjects exhibited very high alpha levels during the recordings they could hardly be asleep or even drowsy. All subjects also stated afterwards that they had not been asleep but had been consciously absorbed in their inner subjective experiences.
Ayahuasca is thought by indigenous people to give the individual access to unconscious, spiritual and transpersonal realms of experience. The psychological state induced by ayahuasca can be compared to other altered states of heightened unconscious activity such as meditation, hypnosis, etc. A number of studies have shown that the presence of theta waves in the EEG reflects subconscious activity, and people with increased EEG theta levels usually have better access to their subconscious feelings, imagery and memories (Green E. & Green A., 1977; Budzynski, T.H., 1986; Cowan, J., 1993; White, N.E., 1999). During states of increased awareness of subconscious activity (e.g. meditation, hypnotic trance, hypnagogic imagery) theta activity usually increases considerably. The presence of alpha waves in these states is important since they reflect conscious attention. If alpha amplitudes drop down it is a sign that the individual is losing consciousness. This is what happens when a person falls asleep.
Budzynski (1986) says "In contrast to the waking or deep sleep states, there is a very special state of light sleep. Identified by an EEG pattern of theta frequencies (4-7Hz), this transitory condition has been referred to as the reverie state, the fringe of consciousness, the pre-conscious, and the twilight state�. During this rather brief period � people often experience emergent, hallucinatory, dreamlike experiences that are more disjointed and brief than those dreams associated with rapid eye movement (REM) sleep� A number of illustrious individuals from the fields of science, music, literature and art have credited the imagery produced during the twilight state for creative solutions or inspiring thoughts." (Budzynski, 1986, pp. 429-30).
Ayahuasca seems to induce and expand the hypnagogic or twilight state (correlated with an increase of theta) and at the same time keeps the individual awake and conscious (correlated with an increase of alpha). These EEG changes are exactly the same as during deep meditation (Hoffmann, E. 1998). Thus, in some respects the ayahuasca-altered state is comparable to meditation and it is quite conceivable that ayahuasca facilitates meditation.
In an altered state of increased EEG theta and alpha activity the individual exhibits greater susceptibility and better conscious access to his unconscious realms of experience without interference from his critical, analytical mind which is silenced. This is exactly what happens in hypnosis and, we believe, during the altered state of consciousness induced by ayahuasca. However, in this latter state the individual is his own hypnotist and therapist. He may have new profound insights into his own behavior patterns and may learn to integrate previously unconscious material at a higher level of consciousness. In neurological terms the individual learns to raise, for example, emotional conflicts from a limbic to a neocortical level where the problems can be processed, understood and integrated.
These speculations are supported by extensive studies of ketamine-assisted psychotherapy done in Russia. Ketamine is an anaesthetic used in surgery which in small doses induces a profound psychedelic experience lasting for about an hour. Krupitsky and Grinenko (1998) reported a 3-4 fold increase of theta activity in all regions of the cortex following the ingestion of ketamine. These authors suggest that their findings of increased theta are "...evidence of limbic system activation during ketamine sessions, as well as evidence for the reinforcement of the limbic-cortex interaction. This fact can also be considered to a certain extent to be indirect evidence for the strengthening of the interactions between the conscious and subconscious levels of the mind during the KPT (Ketamine Psychedelic Therapy)", (Ibid. p. 60).
Our data do not support previous findings of cortical activation with decreased alpha and increased beta activity caused by psychedelics such as LSD, mescaline and psilocybin (Wikler A, 1954; Itil TM, 1968; Fink M, 1978). Neither can the hyperfrontality hypothesis with increased frontocortical activation (Vollenweider F.X., 1998) in psychedelic states be supported. The only significant EEG change we found in the frontal lobes was an increase of theta waves reflecting a state of deep relaxation and reverie. Thus, ayahuasca seems to have different effects on brain functions than the traditional psychedelics (LSD, mescaline, psilocybin). Our findings rather point to a similarity between the altered states produced by ayahuasca and marihuana which both appear to stimulate the brain to produce more alpha waves (Fink M, 1976, Lukas et al., 1995).
Ayahuasca has a long history of use for healing and spiritual purposes in shamanistic rituals in South America (Metzner, R., 1999). Psychological long-term effects of ayahuasca have been investigated recently by Grob et al., (1996) who compared a group of 15 long-term users with 15 controls with no prior use of ayahuasca. The assessment included psychiatric diagnostic interviews, personality and neuropsychological testing. The study reported considerable remission from certain psychopathology and substance abuse among the long-term users with no evidence of personality or cognitive disturbances. Overall assessment revealed a high functional status of ayahuasca users with no safety issues (Grob, C.S. et al., 1996; Grob C.S., 1999). If further studies support the notion that ayahuasca opens up people to their unconscious feelings and memories and gives them an opportunity to explore new psychological insights, ayahuasca might be used as a valuable adjunct to psychotherapy.
References
Budzynsky TH. 1986. Clinical applications of non-drug-induced states. In Handbook of States of Consciousness, Wolman BB, Ullman M (eds.). Van Nostrand Reinhold Co.: New York.
Callaway JC, McKenna DH, Grob CS, Brito GS, Raymon LP, Poland RE, Andrade EN, Andrade EO, Mash DC. Pharmacokinetics of Hoasca alkaloids in healthy humans. J Ethnopharmacol. 1999 Jun;65(3):243-56.
Cowan JD. 1993. Alpha-theta brainwave biofeedback: The many possible theoretical reasons for its success. Biofeedback 21 (2), 11-16.
Dafters RI, Duffy F, O'Donnell PJ, Bouquet C. 1999. Level of use of 3,4-methylenedioxymethamphetamine (MDMA or Ecstacy) in humans correlates with EEG power and coherence. Psychopharmacology (Berl) 145(1): 82-90.
Don NS, McDonough BE, Moura G, Warren CA, Kawanishi K, Tomita H, Tachibana Y, Böhlke M, Farnsworth NR. 1998. Effects of Ayahuasca on the human EEG. Phytomedicine, Vol. 5(2): 87-96.
Fink M. 1976. Effects of acute and chronic inhalation of hashish. Marihuana, and delta 9-tetrahydrocannabinol on brain electrical activity in man: evidence for tissue tolerance. Ann N Y Acad Sci 282: 282-387.
Fink, M. 1978. EEG and psychopharmacology. Contemp. Clin. Neurophys. Suppl. 34: 41-56.
Gamma A, Frei E, Lehmann D, Pascual-Marqui RD, Hell D, Vollenweider FX. 2000. Mood state and brain electric activity in ecstacy users. Neuroreport 11 (1): 157-62.
Green E, Green A. Beyond Biofeedback. Dell Publishing Co.: New York 1977.
Grob CS, McKenna DJ, Callaway JC, Brito GS, Neves ES, Oberlaender G, Saide OL, Labigalini E, Tacla C, Miranda CT, Strassmann RJ, Boone KB. 1996. Human psycho-pharmacology of hoasca, a plant hallucinogen used in ritual context in Brazil. Journal of Nervous and Mental Disease 184: 86-94.
Grob CS. 1999. The psychology of ayahuasca. In: Ayahuasca, Hallucinogens and Consciousness, Metzner R. (ed.). Thunders Mouth: New York, 214-250.
Hoffmann, E. 1998. Mapping the brain's activity after Kriya Yoga. (Scandinavian Yoga and Meditation School. Bindu 12: 10-13.
Itil TM. 1968. Electroencephalography and pharmacopsychiatry. Clin. Psychopharmacol. 1: 163-194.
Krupitsky EM, Grinenko AY. 1997. Ten year study of ketamine psychedelic therapy (KTP) of alcohol dependence. The Heffter Review of Psychedelic Research. Volume I. The Heffter Research Institute, Santa Fe, NM.
Lukas SE, Mendelson JH, Benedikt R. 1995. Electroencephalographic correlates of marihuana-induced euphoria. Drug Alcohol Depend 37 (2): 131-40.
McKenna D, Towers GHN, Abott FS. 1984. Monoamine oxidase inhibitors in South American hallucinogenic plants: Tryptamine and beta-carboline constituents of ayahuasca. Journal of Ethnopharmacology 10:195-223.
McKenna DJ, Grob CS, Callaway JC. 1998. The scientific investigation of Ayahuasca: a review of past and current research. Heffter Review of Psychedelic Research 1: 65-77.
Metzner R (ed.) Ayahuasca: Hallucinogens, Consciousness, and the Spirit of Nature. Thunder's Mouth Press: New York 1999.
Morley BJ, Bradley RJ. 1977. Spectral analysis of mouse EEG after the administration of N,N-dimethyltryptamine. Biol Psychiatry 12(6): 757-69.
Peniston EG, Kulkosky PJ. 1989. Alpha-theta brain wave training and beta-endorphin levels in alcoholics. Alcohol Clin Exp Res 13: 271-79.
Ray WJ. 1997. EEG concomitants of hypnotic susceptibility. Int J Clin Exp Hypn 45 (3): 301-13.
Strassman RJ, Qualls CR, Uhlenhuth EH. 1994. Dose-response study of N,N-dimethyltryptamine in humans. II: Subjective effects and preliminary results of a new rating scale. Archives of General Psychiatry 51: 98-108.
Vollenweider FX. 1998. Recent advances and concepts in the search for biological correlates of hallucinogen-induced altered states of consciousness. The Heffter Review of Psychedelic Research. Volume I. The Heffter Research Institute, Santa Fe, NM.
White NE. 1999. Theories of the effectiveness of alpha-theta training for multiple disorders. In Introduction to Quantative EEG and Neurofeedback. Evans JR, Abarbanel A (eds.). Academic Press: New York.
Wikler A. 1954. Clinical and electroencephalographic studies on the effects of mescaline, N-allylnormorphine and morphine in man. J. Nervous and Mental Dis. 120: 157-175.
Info about the PCEEG8 hard and software may be obtained from Khepri Electronics, Nyvej 17, 3220 Tisvildeleje, Denmark.
 |
 |
 |